· Cultivars · 10 minutes
Anji Bai Cha: on the molecular basis of tea albinism
Anji Bai Cha (安吉白茶) is a modern green tea variety from the Anji region in Zhejiang Province. In the 1970s, an old tea tree with unusually light-colored young shoots was discovered near the village of Dashi. In 1982, work began on vegetative propagation of material from this tree.

As a result of research work, the first batch of tea with the working name Yu Feng (玉凤) — “Jade Phoenix” — was produced in 1991 and was highly rated by experts. By the end of 1990, the study of this variety’s properties and the development of its processing technology were completed, and in 2000 the new cultivar was registered under the name “Bai Ye Yi Hao” (白叶一号) — “white leaf number one,” and the finished product was named Anji Bai Cha. Since 2004, the name Anji Bai Cha has been geographically protected.
Over the past twenty years, the production of Anji Bai Cha has increased many times over and currently amounts to approximately 2,000 tons per year. Anji Bai Cha has become one of the most well-known green teas not only in Zhejiang but also throughout China. Besides Zhejiang, Bai Ye Yi Hao is cultivated in Liyang, Changzhou, Jiangsu, in the Tunzhen autonomous region, in Guizhou Province, and elsewhere. The prices of tea from white-leaf and yellow-leaf cultivars (Bai Ye Yi Hao being the most popular of them, but not the only one) are quite high, and interest in such tea continues to grow, so economically disadvantaged regions see an opportunity to overcome poverty through its production.
Bai Ye Yi Hao is a temperature-sensitive semi-albino. The first buds and leaves have a yellow-green color. At the bud + leaf to bud + 2 leaves stage, at temperatures below 20°C, the leaves acquire a characteristic “jade” color — light green or light yellow, not snow-white but significantly lighter than a “normal” tea bush, while the central vein of the leaf and the areas of lateral veins close to it become dark green, creating a contrasting pattern. With further shoot development, as well as at temperatures above 22°C, the leaves become light green, as is typical for spring tea shoots. For high-quality Anji Bai Cha, the raw material is harvested during the albino stage, so the harvesting window is short — usually about two weeks from mid-March to early April.

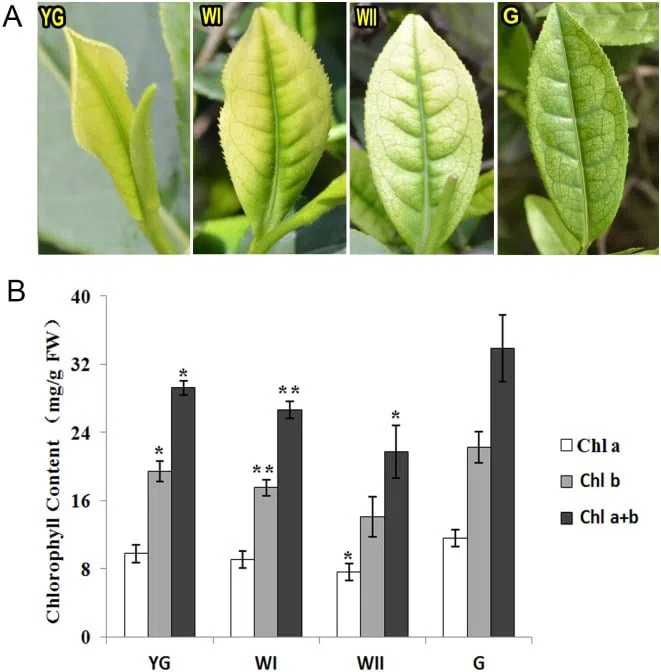
Tea from Bai Ye Yi Hao harvested at the albino stage has an unusual chemical composition. It contains less chlorophyll, which affects the color, but of course this does not affect the taste — chlorophyll is insoluble and contributes almost nothing to the taste and aroma of tea. What is important is something else: Anji Bai Cha contains significantly less caffeine and other purine alkaloids compared to regular green teas; significantly fewer polyphenols — only 10-14%; and significantly more amino acids (primarily theanine, as well as allothreonine, glycine, serine, tryptophan, citrulline, glutamine, alanine, valine, and proline) — up to 6-7%, and in some cases up to 10%, which is many times higher than in regular green teas. This gives Anji Bai Cha a refreshing, sweet taste with a pronounced umami character and minimal astringency and bitterness.
The processes occurring in the leaves of white-leaf and yellow-leaf cultivars during the albino and re-greening stages, along with the mechanisms of their regulation, have attracted considerable scientific attention. The composition of Bai Ye Yi Hao leaves has been studied at the transcriptomic (collection of RNA), proteomic (collection of proteins), and metabolomic (collection of low-molecular-weight endogenous metabolites) levels. Links to the relevant studies are provided at the end; here I want to pause briefly to show just how biochemically complex a factory a tea leaf is.
Chlorophyll is located in thylakoids — membrane structures assembled into stacks called grana inside chloroplasts — the organelles responsible for photosynthesis in plant cells. When chlorophyll biosynthesis decreases — for example, during prolonged darkness — chloroplasts reversibly transform into weakly pigmented etioplasts. At the first “yellow-green” stage, the chloroplasts of Bai Ye Yi Hao had a typical structure with well-defined thylakoids assembled into grana; at the albino stage, the grana disappeared, only a few thylakoids remained in the chloroplasts, and some chloroplasts turned into etioplasts; after the temperature rose during the re-greening stage, the number of chloroplasts increased and their structure returned to normal.
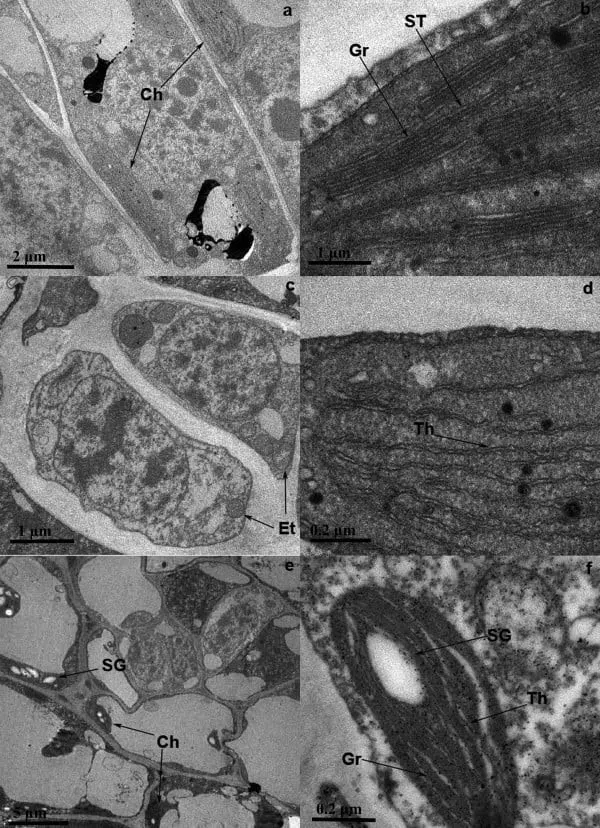
In the proteomic analysis, more than sixty proteins were found whose expression changes significantly depending on the stage of shoot development. These proteins participate in glycolysis and energy metabolism, carbon, nitrogen and sulfur metabolism, photosynthesis, protein and RNA processing, and stress responses. Energy metabolism proteins are represented by phosphoglycerate kinase and enolase. Phosphoglycerate kinase catalyzes the reversible transfer of a phosphate group from 1,3-bisphosphoglycerate to ADP, producing 3-phosphoglycerate and ATP. The expression of this enzyme increases at the albino stage, which may indicate increased energy production for various defensive processes associated with cold stress. The expression of enolase, which catalyzes the conversion of 2-phosphoglycerate to phosphoenolpyruvate, was reduced, but its activity was maintained — it appears that enolase activity is controlled at the post-transcriptional level so that the intensity of glucose oxidation does not decrease at low temperatures.
Proteins involved in nitrogen and sulfur metabolism were identified as S-adenosylmethionine synthase, glutamine synthetase, and methylenetetrahydrofolate reductase; their levels were reduced at the “white” stage. S-adenosylmethionine synthase catalyzes the formation of S-adenosylmethionine from L-methionine and ATP. S-adenosylmethionine acts as a universal methyl (-CH3) donor in many biological reactions. In particular, it is the methyl donor in the formation of protochlorophyll from Mg-protoporphyrin, and protochlorophyll is, in turn, a direct precursor of chlorophyll — its deficiency is one of the causes of leaf albinism. S-adenosylmethionine is also a methyl donor in caffeine biosynthesis, and its shortage is one of the reasons for the low caffeine content in Anji Bai Cha; as we can see, different compounds such as caffeine and chlorophyll share common key points in metabolism. Methylenetetrahydrofolate reductase is an important enzyme in methionine metabolism, and methionine is required in the initial stages of caffeine biosynthesis. Glutamine synthetase catalyzes the formation of glutamine from glutamic acid and ammonia; this enzyme is important for protecting the plant against excess ammonium. In the absence of glutamine synthetase, glutamic acid can be directed toward theanine synthesis, in which excess nitrogen accumulates. From a proteomic point of view, this is a logical assumption, but metabolomic analysis does not confirm it: the glutamine content at the albino stage actually increases and can be converted to glutamic acid and alanine; during the decarboxylation of the latter, ethylamine is formed. Glutamic acid and ethylamine are needed for theanine synthesis, which you will be able to read about in the next part.
Ribulose-1,5-bisphosphate carboxylase (RuBisCO) is the most important enzyme in plant leaves (it is the most abundant enzyme in the world), directly responsible for carbon dioxide fixation in photosynthesis; one could say that it is RuBisCO that performs the transition from inorganic to organic matter. At the albino stage, the content of intact large subunits of RuBisCO decreases while the content of their fragments increases. Reduced photosynthetic activity leads to lower carbohydrate production. Fructose and glucose-1-phosphate levels are reduced during the “white stage,” while glucose levels are reduced at the beginning of the “white” stage and increase toward its end, which appears to reflect increased hydrolysis of reserve polysaccharides. Carbohydrate deficiency leads to the activation of alternative energy-producing pathways. As a result, most phosphoenolpyruvate is directed to the citric acid cycle and oxidized in it, while less goes to the shikimic acid pathway. This explains the reduction in polyphenol production — the pyruvate needed for their synthesis is “burned.”
It should also be taken into account that glutamic acid is consumed in chlorophyll biosynthesis — it stands at the very beginning of the chain of twenty reactions leading to the formation of chlorophyll. And with a decrease in chlorophyll synthesis, there may be an excess of glutamate that can be directed toward theanine synthesis. At the albino stage, the expression of heat shock proteins HSP60 and HSP70 decreases, and HSP70 is essential for proper chloroplast development. Eukaryotic initiation factor 4a (eIF-4A) plays an important role in translation initiation in eukaryotes; a decrease in its content at the albino stage can lead to impaired biosynthesis of new proteins — including the enzymes mentioned above.
Early light-inducible protein (ELIP), a protein from the chlorophyll-binding family, participates in the assembly or repair of the photosynthetic apparatus during early chloroplast development. Its expression is induced by many stress factors. Disruption of chloroplast structure at the albino stage leads to reduced photosynthetic capacity, and under these conditions excess light energy can lead to the formation of toxic compounds — in particular, free radicals. The accumulation of ELIP may protect the tea plant from their damaging effects. Isoflavone reductase participates in the synthesis of isoflavone phytoalexins. Due to reduced polyphenol biosynthesis, isoflavone phytoalexin production is also reduced, so the expression level of isoflavone reductase at the “white” stage is decreased — it is simply not needed.
And this is only part of the discovered differences. The regulation of the described processes is highly complex: 8,790 genes were identified whose expression changes after albinism onset in Bai Ye Yi Hao shoots, and 9,476 genes whose expression changes after re-greening. Transcriptomic analysis not only confirms the conclusions from proteomic and metabolomic studies but also complements them. Thus, notable differences were revealed in carotenoid content at different stages. Carotenoids are terpenoids derived from the acyclic hydrocarbon lycopene; they are pigments of yellow, orange, and red color; they participate in photosynthesis and also serve as precursors of abscisic acid — an important plant hormone. Lycopene content decreased with shoot development, lutein content increased, and beta-carotene content reached its maximum at the “white” stage and decreased after re-greening. This is due to different degrees of expression of genes encoding lycopene beta-cyclase (which converts lycopene to gamma-carotene, from which beta-carotene and zeaxanthin are subsequently formed) and lycopene epsilon-cyclase (which converts lycopene to delta-carotene, from which alpha-carotene and lutein are subsequently formed). For the taste and aroma of green tea, carotenoids are not very important, but during oxidative fermentation in black tea production, carotenoids break down to form terpene compounds that influence the taste and aroma of the tea. Thus, with the development of a Bai Ye Yi Hao shoot, there is not only a decrease and then restoration of chlorophyll synthesis — leading to changes in leaf color — but also a profound rearrangement of metabolic pathways; upon close examination, these reveal connections between seemingly unrelated substances such as chlorophyll, alkaloids, polyphenols, and theanine. Why would I want to write this article that is two-thirds advanced biochemical knowledge, not understandable to some readers?
First, no less complex processes occur during the development of any green tea shoots. In white-leaf and yellow-leaf cultivars, these changes are more pronounced, but in ordinary tea the content of caffeine, amino acids, and polyphenols also changes depending on both the stage of shoot development and external conditions. The chemical composition of tea raw material from the same garden, harvested at different times, can differ no less than the composition of raw material from different cultivars and different locations. Therefore, the interest in tea harvested at a specific optimal time for that cultivar, and the high price of such tea, have quite objective reasons. In many green tea cultivars, early tea is highly valued, and this is not about tradition, not about marketing, not about the long wait for this tea to appear. It actually — biochemically! — differs from tea of later harvests. Anji Bai Cha is a perfect example of this.
Second, I wanted to remind that besides the world in which people prefer not to seek explanations but to attribute everything to mystical, fundamentally unknowable “energies,” there is another world — a world of scientific inquiry, intelligence, ingenuity, and audacity. And the achievements of this world are impressive.
Bibliography: - 1, 2, 3, 4 , 5
Transcriptomic analysis Proteomic analysis Metabolomic analysis
Source: Tea shop “Owl and Panda”



